


Аррения эпихизиевидная (Arrhenia epichysium)
- Отдел: Basidiomycota (Базидиомицеты)
- Подотдел: Agaricomycotina (Агарикомицеты)
- Класс: Agaricomycetes (Агарикомицеты)
- Подкласс: Agaricomycetidae (Агарикомицетовые)
- Порядок: Agaricales (Агариковые или Пластинчатые)
- Семейство: Hygrophoraceae (Гигрофоровые)
- Род: Arrhenia (Аррения)
- Вид: Arrhenia epichysium (Аррения эпихизиевидная)
Синонимы:
-
Омфалина кубковидная
- Омфалина эпихизиевидная
- Клитоцибе эпихизиевидная
- Плевротеллус эпихизиевый
- Аррения кубковидная

Полное научное название: Arrhenia epichysium (Pers.) Redhead, Lutzoni, Moncalvo & Vilgalys
Этимология названия
— Arrhenia: в честь шведского миколога Й. Аррениуса (J. Arrhenius).
— Epichysium: от греческого epi (на) + chrysos (χρυσός) (золото) – вероятно, отсылка к желтоватому оттенку пластинок у старых грибов.
История систематики
Вид был впервые описан Христианом Персоном в 1801 году как Merulius epichysius. Долгое время его относили к роду Omphalina из-за характерной воронковидной шляпки. Род Omphalina долгое время был искусственной группой (полифилетическим таксоном) куда включали виды сходные по морфологии, но генетически разнородные.
Переломный момент наступил в 2002-2008 годах, когда молекулярные исследования (Redhead et al.) показали его близкое родство с лихенизированными видами рода Arrhenia. Ключевые доказательства:
- Генетические маркеры (ITS и LSU рДНК)
- Наличие симбиоза с водорослями Coccomyxa
- Особенности микроструктуры (пряжки на гифах, форма цистид)
Официальный перенос в род Arrhenia зарегистрирован в Index Fungorum в 2008 году. В русскоязычной литературе старое название Омфалина эпихизиевидная, продолжало использоваться до 2010-х годов, но после генетических исследований так же перешли на Аррения.
Этот скромный грибок – живое свидетельство того, как современная наука переписывает систематику, опираясь на генетику, а не только на внешние признаки.
Описание
Шляпка: у молодых экземпляров шляпка выпуклая с подвёрнутым краем, затем становится распростёртой или даже воронковидной с характерным углублением в центре. Диаметр 0,5–3 см (обычно около 1–1,5 см). Край часто волнистый, у старых грибов может растрескиваться.
Поверхность шляпки:
В сырую погоду – гладкая, слегка клейкая, тёмно-серая или серо-коричневая с оливковым оттенком
В сухую – становится матовой, бледнеет до серовато-бежевого цвета.

Мякоть тонкая (1–2 мм), гигрофанная. После дождя шляпки становятся полупрозрачными, хорошо видны радиальные полоски – следы пластинок с нижней стороны.

Пластинки: редкие (12–15 на шляпку), широкие (до 3 мм), нисходящие по ножке. Между основными пластинками часто встречаются короткие промежуточные.
Цвет пластин: у молодых грибов беловатые, затем приобретают сероватый или желтоватый оттенок.

Ножка: цилиндрическая, иногда слегка изогнутая, 1–4 см в высоту, 1–3 мм в толщину. Обычно немного светлее шляпки, в верхней части может быть почти белой. Поверхность продольно-волокнистая, у основания иногда с беловатым опушением. Мякоть жёсткая, волокнистая.
Ножка часто врастает в субстрат, и при сборе гриб обычно выдёргивается с кусочком древесины или мха.

Запах и вкус:
Слабый, едва уловимый запах. Описывается как нейтральный, слегка «грибной», иногда с лёгким землистым или сырым оттенком.
Мякоть почти безвкусная. Нет выраженной горечи, остроты или сладости. После разжёвывания остаётся лёгкое древесно-травянистое послевкусие, иногда едва заметная пресная водянистость.
Микроскопия:
Споры 7–10 × 4–6 мкм, эллипсоидные, иногда слегка миндалевидные, абсолютно гладкие. Слабо амилоидные (в реактиве Мельцера голубеют).
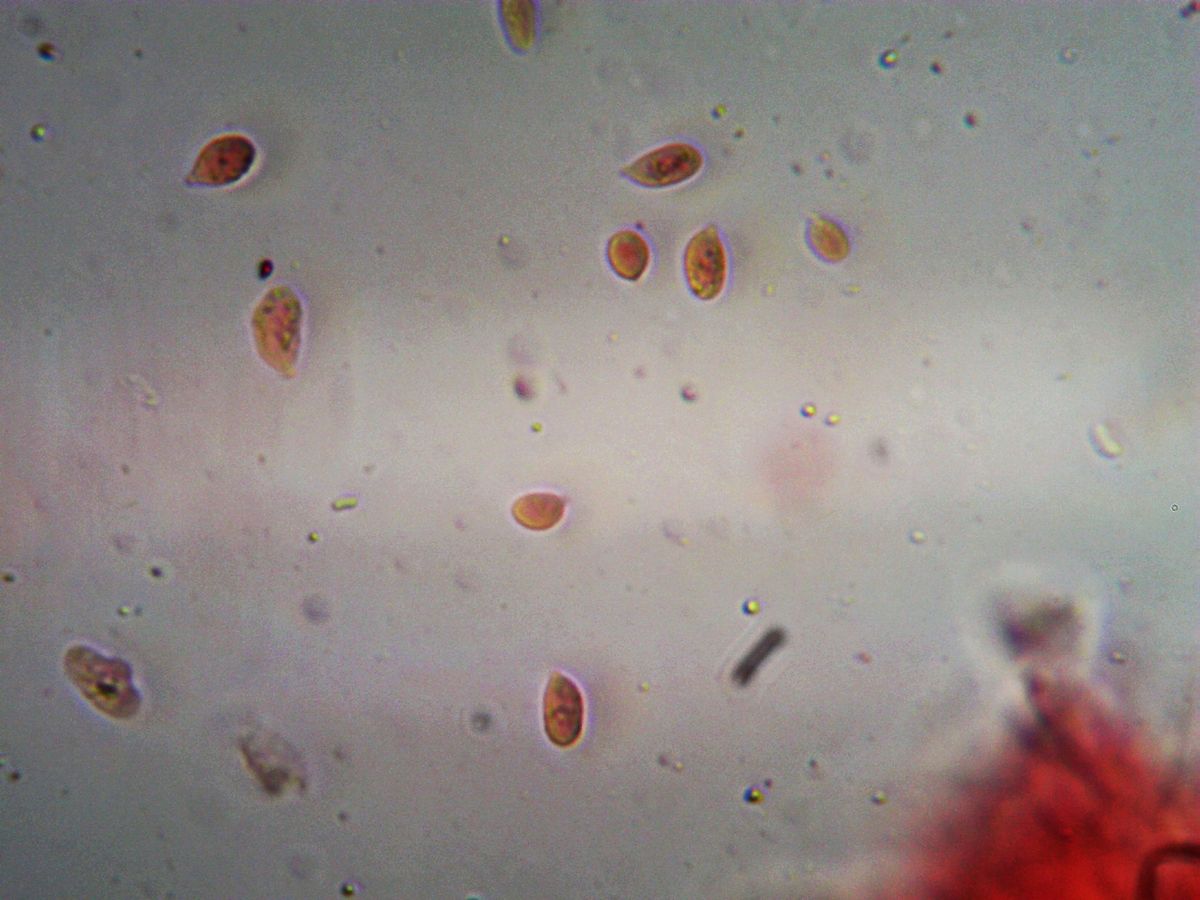

Базидии 25–35 × 5–7 мкм. Булавовидные, всегда 4-споровые, с пряжкой.
Хейлоцистиды многочисленные, булавовидные или веретеновидные, 30–50 × 5–10 мкм.
Плевроцистиды отсутствуют.
Гифальная система мономитическая, гифы с пряжками.
Под микроскопом особенно красивы хейлоцистиды – они образуют «бахрому» по краю пластинок.
Особенности лихенизации
Данный гриб входит в немногочисленный клуб базидиолишайнков. Однако гриб способен и к сапротрофному питанию, разлагая древесину, и может существовать без водорослей. Такой тип взаимоотношений называется — факультативный симбиоз.
Фотобионтом грибу служат водоросли Coccomyxa.
Водоросли локализуются в гименофоре (пластинках) и под кутикулой шляпки. Образуют точечные скопления между гифами, но не сплошной слой. В мицелии водоросли встречаются фрагментарно в наружных частях мицелия. Нет чёткой зональности, как у настоящих лишайников.
Гифы образуют рыхлую сеть вокруг фотобионта. Клетки Coccomyxa диаметром 5–10 мкм. Располагаются поодиночке, не образуют кластеров.
Настоящий таллом у Аррении эпихизиевидной отсутствует. Нет дифференцированных слоёв (коры, сердцевины и т.д.). Нет специализированных структур для удержания водорослей (гаусторий, цист). Гифы лишь оплетают клетки водорослей, не проникая внутрь.
При недостатке света гриб усиливает лихенизацию. Например в тёмных ельниках, где мало света, лихенизация выражена сильнее – гриб буквально «цепляется» за возможность фотосинтеза.
В сухих условиях водоросли переходят в покоящееся состояние.
Связь с мхами
В данном случае речь идёт о тройном взаимодействии в котором задействованы
- Гриб (Arrhenia epichysium)
- Водоросли (Coccomyxa)
- Мхи (преимущественно Polytrichum alpinum, Racomitrium lanuginosum)
Аррения эпихизиевидная практикует два вида взаимодействия:
Эпифитный вариант:
Гриб растёт на живых мхах, образуя связь со мхом, при которой гифы проникают между листочками мха.
Подстилочный вариант:
В местах, где мох образует плотный ковёр, гриб использует отмершие части мхов как субстрат, при этом гифы гриба формируют «мостики» между древесиной и моховым слоем.
Хотя этот гриб в первую очередь образует симбиоз с водорослями, его взаимодействие с мхами (особенно в альпийских и северных регионах) даёт несколько ключевых преимуществ:
- Укрытие от пересыхания. Моховые «подушки» (Polytrichum, Racomitrium) сохраняют влажность до 95% даже в засуху, что критично для тонких гиф.
- Защита от ультрафиолетового излучения. В высокогорьях мох снижает дозу ультрафиолета на 40-60%.
- Стабильность температуры. Под мхом суточные колебания меньше на 5-8°C.
- Доступ к питательным веществам. Мхи (особенно Sphagnum) выделяют до 0.3 мг/л аммония при разложении – это важный источник азота в бедных субстратах.
- Минеральный обмен. Гифы гриба проникают в отмершие части мхов, получая калий и фосфор (исследования трансмиссионной электронной микроскопии показали перенос через гифальные «мостики»).
В числе дополнительных бонусов для гриба от такой связи можно назвать, например, улучшенное закрепление. На гнилой древесине гриб часто слабо прикреплён, а ризоиды мхов служат «якорями» для мицелия.
Например, в Пиренеях 78% находок растут именно в местах контакта мха и древесины.
Для гриба мох – это «живой инкубатор», улучшающий выживаемость в экстремальных условиях. Настоящий симбиоз здесь только с водорослями, а связь с мхами – скорее комменсализм.
Безусловно симбиоз Аррении и мхов не равноценен – гриб получает больше выгод. Всё же и для мхов польза есть. Например, гифы ускоряют разложение отмерших частей мха, высвобождая минералы, а так же мицелий защищает от некоторых патогенных грибов (например, Pythium).
Экология
Основной субстрат: гнилая древесина хвойных пород (особенно ели и сосны), разлагающиеся пни, покрытые мхом валежины. Иногда может вырастать на плотном слое лесной подстилки.
Аррения эпихизиевидная никогда не образует крупных скоплений. Чаще всего плодовые тела появляются одиночно или небольшими группами по 3–5 экземпляров.
Встречается Аррения в ельниках (особенно черничных и кисличных), в сосняках-зеленомошниках, в старых пихтовых лесах. Кроме того, увидеть её можно на обочинах старых лесных дорог с брошенной гниющей древесиной и заброшенных лесопилках (на остатках хвойных брёвен).
Предпочитает затенённые участки с высокой влажностью, близ ручьёв, лесных болот, а так же северные склоны в горных районах.
Сезон плодоношения
В среднем, временем плодоношения этого гриба считается сезон с июля по ноябрь. В разных регионах сезон может смещаться.
Например, в Северной Европе (Скандинавия, Карелия, Архангельская обл.) сезон короткий и интенсивный с середины августа до конца сентября. В Подмосковье и Ленинградской области сезон плодоношения стабильный, продолжительный. Сроки с конца августа до середины октября. А в Южных регионах (Кавказ, Крым) сезон длинный и смещённый – с июля по ноябрь.
Географическое распространение
Основной ареал Аррении эпихизиевидной сосредоточен в Европе, Северной Азии и Северной Америке.
В России встречается по всей лесной зоне, включая Дальний Восток. В Европе от Скандинавии до Балкан и Кавказа. В Южной Европе очаги обитания этого вида сосредоточены в горных районах Альп и Пиреней.
В Северной Америке встречается от Аляски до Великих озёр.
Не растёт на лиственных породах в тропиках.
Этот гриб нельзя назвать ни распространённым, ни крайне редким. Многое сильно зависит от региона и условий.
Съедобность
В старых определителях иногда указывался как «несъедобный», но не ядовитый. Современные микологи (например, по данным [Микологического общества РАН, 2020]) вообще не рассматривают его как пищевой объект, поэтому токсичность гриба изучена недостаточно.
Сходные виды

Лихеномфалия зонтиконосная (Lichenomphalia umbellifera)
Ещё один вид лихенизированного базидиомицета. Это их объединяет. Плодовые тела по внешнему виду, так же схожи. Это мелкие пластинчатые грибы с воронковидный шляпкой и нисходящими пластинками.Давайте разберемся в различиях. Прежде всего стоит обратить внимание на наличие таллома. Не лихенизированные плодовые тела Лихеномфалии зонтиконосной вырастают из зелёного, мелкозернистого, желеобразного таллома. У Аррении эпихизиевидной всё наоборот. Основная лихенизация сосредоточена в плодовых телах, а ничего подобного на таллом не наблюдается.
Оба вида растут на мёртвой древесине хвойных пород, Лихеномфалия может жить и на почве, камнях или гниющем мхе. Тогда как Аррения эпихизиевидная связана исключительно с хвойной древесиной.
Если говорить о внешнем облике плодовых тел, то шляпка Лихеномфалии чаще светло-коричневая и не гигрофанная, тогда как у Аррении эпихизиевидной она серо-оливковая и гигрофанная.
Различия в микроскопии: споры Лихеномфалии крупнее (9–12 × 5–7 мкм) и не амилоидные, а у Аррении эпихизиевидной споры мельче (7–10 × 4–6 мкм) и слабо амилоидные.
Экологически Лихеномфалия тяготеет к открытым влажным местообитаниям (тундры, горные луга), тогда как Аррения предпочитает тенистые хвойные леса.
Аррения тёмная (Arrhenia obscurata)
Это ещё один вид базидиолишайника с мелкими плодовыми телами (шляпка 0.5–3 см) и нисходящими пластинками. Всё это, без условно делает эти два вида схожими визуально. Однако и различий хватает.
Первое это более тёмная, почти чёрная шляпка у Армении тёмной, тогда как у Аррении эпихизиевидной она серо-коричневая. Второе отличие это субстрат. Arrhenia obscurata живёт на древесине лиственных пород (берёза, ольха). В отсутствии таковой, например в тундровой зоне и высокогорье может встречается на песчано-каменистых субстратах, покрытых мхами. Arrhenia epichysium живёт на древесине хвойных пород.
Кроме того у Аррении тёмной споры чуть крупнее (8–11 × 5–6 мкм).
Омфалина бледно-серая (Omphalina griseopallida)
Опять таки сходство во внешнем виде плодовых тел. У обоих видов они мелкие с воронковидными шляпками и нисходящими пластинками. И оба растут на мертвой древесине.
Теперь о различиях. Прежде всего Omphalina griseopallida отсутствует лихенизация.
Визуально их отличают пластинки. У Омфалины бледно-серой они узкие и частые, а у Аррении эпихизиевидной пластики широкие и редкие.
Омфалина бледно-серая растет как на хвойных, так и на лиственных субстратах. Для Аррении эпихизиевидной характерна только хвойная древесина.
Микроскопия: У Omphalina griseopallida цистиды отсутствуют.
Дополнительный список синонимов:
- Omphalina epichysium (Pers.) Quél. (1886)
- Merulius epichysius Pers. (1801) – базионим
- Clitocybe epichysium (Pers.) P. Kumm. (1871)
- Pleurotellus epichysius (Pers.) Konrad & Maubl. (1924)
- Sooty Navel
- Lichen Omphalina



















Отвал башки. Гриб-лишайник???