


Ксантория настенная (Xanthoria parietina)
- Отдел: Ascomycota (Аскомицеты)
- Подотдел: Pezizomycotina (Пезизомикотины)
- Класс: Lecanoromycetes (Леканоромицеты)
- Подкласс: Lecanoromycetidae (Леканоромицетовые)
- Порядок: Teloschistales (Телосхистовые)
- Подпорядок: Teloschistaceae (Телосхисты)
- Семейство: Teloschistaceae (Телосхистовые)
- Подсемейство: Xanthorioideae (Ксанториевые)
- Род: Xanthoria (Ксантория)
- Вид: Xanthoria parietina (Ксантория настенная)
Синонимы:
- Lichen parietinus L. (1753)
- Parmelia parietina (L.) Ach. (1803)
- Physcia parietina (L.) DC. (1805)
- Teloschistes parietinus (L.) Norm. (1852)
- Placodium parietinum (L.) Hepp (1853)

Научное название: Xanthoria parietina (L.) Th. Fr. (1860)
Общие названия
- Common orange lichen (Обыкновенный оранжевый лишайник)
- Yellow scale (Жёлтая чешуйка)
- Maritime sunburst lichen (Морской солнечный лишайник)
- Shore lichen (Прибрежный лишайник)
- Golden shield lichen (Золотой щитовой лишайник)
История изучения Ксантории
Ещё до научной систематики люди замечали яркий лишайник на стенах церквей и замков. В средневековых европейских травниках XV–XVI веков его называли «Gelber Mauerschwamm» («жёлтая настенная губка»), подчёркивая характерный субстрат и цвет.
Переломный момент наступил в 1753 году, когда Карл Линней в своём труде Species Plantarum официально описал вид под именем Lichen parietinus. Учёный указал: «Habitat in Europae muris» («растёт на европейских стенах»), закрепив связь организма с антропогенными постройками. Однако в его концепцию вида случайно попали и другие оранжевые лишайники, например Rusavskia elegans, что позже вызвало путаницу.
XIX век стал эпохой таксономических поисков. В 1803 году «отец лихенологии» Эрик Ахариус перенёс вид в род Пармелия (Parmelia), а всего два года спустя Огюстен Декандоль отнёс его к Фисция (Physcia). В 1852 году Йоханнес Норман, очарованный сходством с Teloschistes flavicans, ошибочно записал вид как Teloschistes parietinus. Лишь в 1860 году шведский миколог Теодор Фриз исправил эту ошибку, выделив новый род Xanthoria и закрепив за видом имя Xanthoria parietina. В качестве типового образца он выбрал иллюстрацию из книги Диллениуса Historia Muscorum (1742), изображающую характерное слоевище.
XX век начался с детективной истории: при ревизии гербария Линнея обнаружилось, что под именем Lichen parietinus хранятся пять разных образцов. Потребовались десятилетия споров, чтобы в 1984 году утвердить неотип — чётко идентифицированный образец из Швеции (UPS Herbarium). Параллельно в 1960-х биохимики совершили прорыв, выделив пигмент париетин. Этот антрахинон не только давал лишайнику оранжевый цвет, но и служил щитом от ультрафиолета, объясняя невероятную выживаемость вида.
На рубеже XXI века филогенетические исследования переписали историю вида. ДНК-анализ (ITS, mtSSU) показал, что многие «родственники» Xanthoria parietina на деле являются самостоятельными видами. Например, Xanthoria aureola (когда-то считавшаяся его подвидом) генетически обособилась, адаптировавшись к арктическим условиям. А вот Xanthoria coomae и Xanthoria polessica, описанные в конце 1990-х, после перепроверки в 2020 году оказались лишь морфовариантами parietina.

Морфология и анатомия
Таллом Ксантории листоватый, состоит из округлых, слегка волнистых лопастей, собранных в розетковидные структуры. Крепится к субстрату ризинами (нитевидными выростами на нижней поверхности), не срастаясь с ним полностью. Такой тип таллома относится к морфологической группе — Пармелоидные лишайники (Parmelioid lichens).
Листоватые розетки таллома диаметром от 2 до 20 см, состоящие из радиально расходящихся лопастей. Каждая лопасть имеет округлую или слабо лопастную форму с волнистыми, иногда надрезанными краями, достигая ширины 1–7 мм. Молодые лопасти часто выпуклые и напоминают крошечные купола, тогда как старые становятся плоскими и могут накладываться друг на друга. Центр тесно прижат к субстрату, обычно в виде цельной плёнки. Всё слоевище крепится к субстрату ризинами (нитевидными выростами на нижней поверхности), не срастаясь с ним полностью.
Xanthoria parietina — чемпион изменчивости среди листоватых лишайников. За 300 лет исследований описано свыше 40 морфологических форм, но все они — лишь крайние точки единого континуума адаптаций. Вид мастерски «настраивает» свой облик под среду:
- — В смоговых городах его лопасти утолщаются, превращаясь в биофильтры для тяжёлых металлов;
- — На альпийских скалах он «съёживается» в миниатюрные жёлтые розетки, выдерживая ледяные ветра;
- — В сырых дубравах покрывается выпуклыми «пузырями», усиливая газообмен;
- — Под пологом леса теряет оранжевый пигмент, маскируясь под зелёные водоросли.
Эта пластичность — не хаос, а гениальная стратегия. Один геном — бесконечные фенотипы. ДНК-анализы подтверждают, что это всё формы одного вида, а их различия управляются «включением/выключением» генов в ответ на стресс.
Верхняя поверхность лопастей – наиболее заметная часть таллома. Её яркая жёлто-оранжевая окраска под прямым солнцем служит визитной карточкой вида. Этот «солнечный загар» создаётся пигментом париетином, выполняющим роль УФ-фильтра. В тени или у старых экземпляров центр розетки приобретает оливково-зелёный оттенок (проявляется хлорофилл водорослей). Поверхность гладкая, без вегетативных выростов (соредий/изидий), но края лопастей могут покрываться лёгким беловатым налётом (пруиной).

Нижняя поверхность образует резкий контраст с ярким верхом. Её основной фон – беловатый, кремовый или бледно-серый. Эта поверхность густо усеяна ризинами – нитевидными разветвлёнными выростами длиной 0,5–2 мм, предназначенными для крепления к субстрату. Ризины рыжеватые, оранжевые или светло-коричневые, создают характерный эффект «веснушчатости» на светлом фоне. Текстура нижней стороны – бархатисто-ворсистая, что особенно заметно при боковом освещении.
Анатомически ксантория — гетеромерный организм. На поперечном срезе (толщиной 250–600 мкм) видны четыре зоны:
- — Верхняя кора (с пигментом париетином),
- — Непрерывный слой водоросли Trebouxia,
- — Рыхлая сердцевина для хранения влаги,
- — Нижняя кора с ризинами
1. Верхняя кора (15–30 мкм) — плотный защитный слой из желатинизированных гиф гриба толщиной 3–5 мкм, плотно переплетенных в параплектенхиму (псевдопаренхиму). Гифы пропитаны кристаллами париетина размером 0.5–2 мкм, придающими оранжевый цвет и блокирующими до 95% УФ-излучения.
2. Водорослевый слой (50–120 мкм) — непрерывный пласт клеток зелёной водоросли Trebouxia диаметром 10–15 мкм. Гифы гриба оплетают их, формируя гаустории, которые проникают в клеточные стенки для обмена сахарами и минералами.
4. Сердцевина (150–400 мкм) — рыхлая «губка» из неразветвлённых гиф тоньше (2–3 мкм), ориентированных перпендикулярно субстрату. Межклеточные полости (10–30 мкм в диаметре) запасают воду (до 300% от сухой массы!), позволяя лишайнику переживать засуху.
5. Нижняя кора (10–25 мкм) — тонкий слой с ризинами. Пучки гиф здесь пропитаны лишайниковыми кислотами, защищающими от микробов и высыхания.
Репродуктивные органы
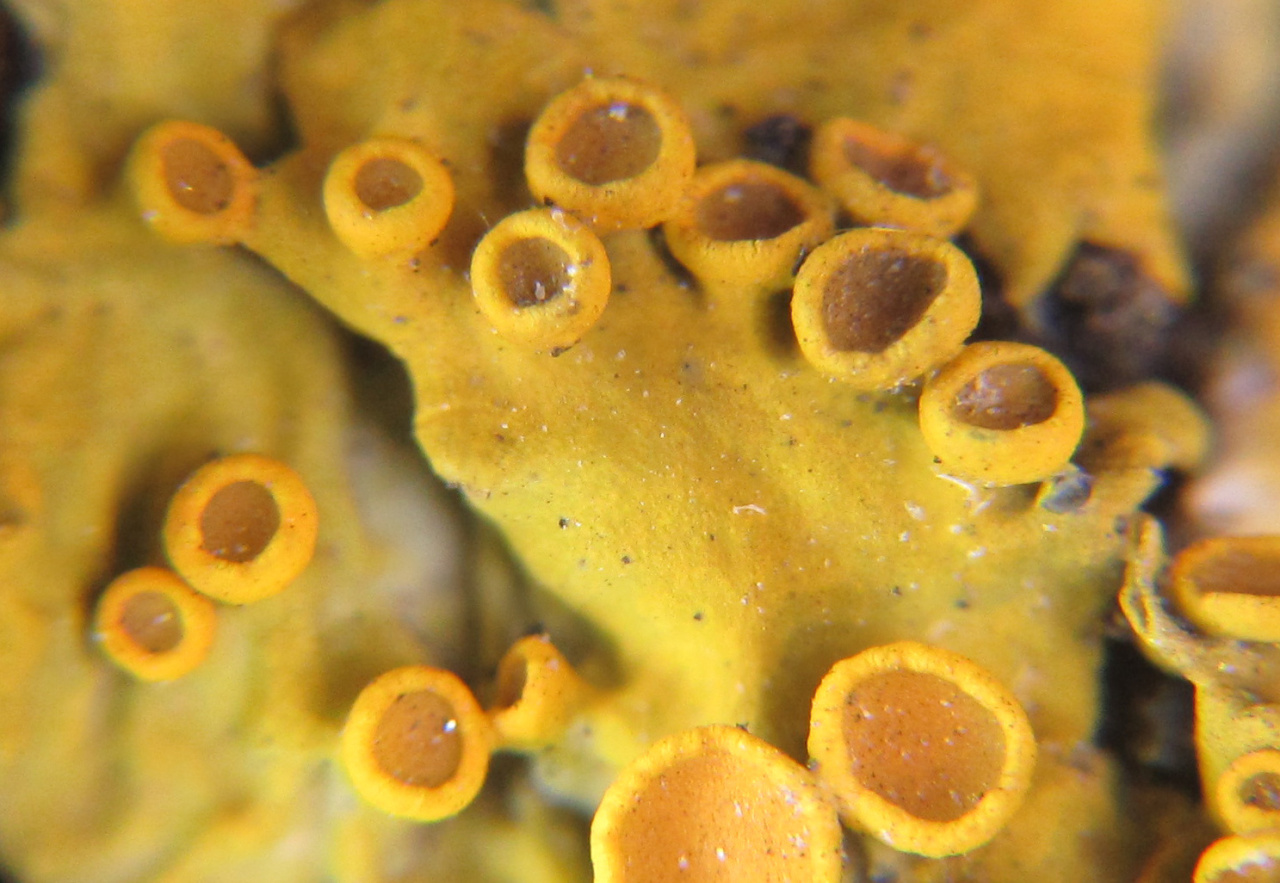
В центре розеток развиваются апотеции — плодовые тела для полового размножения. Они выглядят как миниатюрные (1–5 мм в диаметре) оранжевые «блюдца» с ровным краем. Их диск открыт солнцу и ветру, что характерно для гимнокарповых лишайников. При микроскопии видно, что в апотециях формируются сумки (аски) с восемью двухклеточными спорами каждая.

В продольном разрезе апотеций ксантории напоминает многослойный «торт»:
Сверху расположен эпитеций — тонкий (15–25 мкм) оранжевый слой из пигментированных гиф, насыщенных париетином. Под ним лежит «начинка» — гимений толщиной 250–350 мкм, где вертикально упакованы Аски (булавовидные сумки по 8 спор каждая) и нитевидные парафизы, разделяющие их. Основанием служит гипотеций — плотное переплетение гиф, проводящее питательные вещества из таллома. Уникальная черта — слоевищный край: бордюр из гиф и клеток водоросли Trebouxia, окружающий диск. Этот «живой ободок» не только защищает молодые споры, но и ведёт фотосинтез, пока апотеций созревает. Такая структура обеспечивает выброс спор на расстояние до 60 см при созревании!
Для бесполого размножения служат пикнидии — микроскопические чёрные точки (0.1–0.3 мм), рассеянные по краям лопастей. Их стенки сложены из тёмно-коричневых гиф с толщиной стенок 3–4 мкм. Они производят конидии — клетки размером 4–6 × 1 мкм, которые разносятся ветром и дают начало новым талломам.
Ксантория настенная не образует вегетативных пропагул (соредий или изидий).
Микроскопия
Аски (сумки) булавовидные, размером 50–65 × 12–16 мкм, содержат по 8 спор.
Споры бесцветные, эллипсоидные, одноклеточные, размером 12–16 × 5–9 мкм.
Парафизы нитевидные гифы толщиной 1.5–2 мкм с вздутыми кончиками (до 4 мкм), неокрашенные.
Конидии прямые или слабо изогнутые палочки размером 4–6 × 1 мкм.
Симбиоз с водорослями
Ксантория настенная представляет собой классический хлоролишайник — организм, чей фотосинтез обеспечивается исключительно зелёными водорослями. Её единственным фотобионтом служит Trebouxia (Требуксия). Этот фикобионт — один из самых распространённых в мире лишайников, встречающийся у 70% видов.
Клетки Trebouxia округлые или эллипсоидные, диаметром 10–15 мкм, с гладкой оболочкой. Хлоропласты чашевидные, зелёные, заполняющие 60–80% объёма. Эти водоросли образуют непрерывный слой под верхней корой таллома, где их окружают гифы гриба, формирующие гаустории (специальные выросты для питания).
Современные исследования (2020-е гг.) трактуют отношения гриба и водоросли как «контролируемый паразитизм», а не равноправный мутуализм. Другие источники их взаимоотношения описывают как «облигатный симбиоз с принуждением». Гриб ведёт себя как «фермер», эксплуатирующий водоросль, но обеспечивающий её выживание в агрессивной среде. При нарушении баланса (например, при дефиците света) гриб может уничтожить часть клеток водоросли.
В любом случае гриб доминирует, проникая гаусториями в клеточные стенки Trebouxia и забирая до 80% продуктов фотосинтеза — глюкозу, эритрит и рибитол (сахароспирт), который служит основным источником питания для гриба. Взамен водоросль получает минеральные соли, защиту от УФ через пигмент париетин и концентрированный CO₂.
Гриб подавляет деление клеток водоросли, блокируя их размножение внутри таллома. Для этого гифы выделяют ингибиторы митоза (белки-супрессоры деления клеток). Это позволяет перенаправить ресурсы водоросли на производство сахаров. Если в лаборатории удалить гриб, Trebouxia начинает делиться через 48 часов, но в природе её конкурентоспособность уже в 3 раза ниже свободноживущих штаммов.
Обновление водорослей происходит не за счёт деления, а через захват из окружающей среды. Молодые гифы гриба захватывают клетки Trebouxia из воздуха (концентрация: 200–500 кл/м³) благодаря белкам-аттрактантам. Для покрытия 1 см² нового таллома достаточно 50–100 клеток — эквивалент фильтрации 0.3 л воздуха.
Гриб безошибочно идентифицирует «свою» Trebouxia через три уровня проверки:
1. Рецепторы-антенны на кончиках гиф (белки LSRK) связываются с уникальными углеводами клеточной стенки водоросли.
2. Химический тест: Гриб выделяет оксид азота (NO). Совместимые штаммы нейтрализуют его глутатионом, несовместимые — погибают.
3. Генетическая синхронизация: У партнёров есть общие гены (например, ftsZ для регуляции деления).
Уникальная черта симбиоза — «хамелеон-эффект», это способность ксантории менять цвет при увлажнении — от ярко-оранжевого до зелёного. В сухом состоянии сжатые клетки Trebouxia и плотный слой пигмента париетина в верхней коре работают как оптический фильтр: париетин поглощает сине-зелёные лучи спектра, оставляя для отражения только жёлто-оранжевые волны. Одновременно сплющенные хлоропласты водорослей минимизируют видимость зелёного пигмента.
При увлажнении запускается каскад изменений:
В течение 5–10 минут париетин растворяется в воде, превращаясь из оранжевого экрана в полупрозрачную субстанцию.
Клетки Trebouxia набухают, их хлоропласты расправляются и начинают флуоресцировать зелёным светом.
Вода в сердцевине действует как биологическая линза, усиливая и рассеивая зелёное свечение хлорофилла.
Гриб временно ослабляет УФ-защиту, позволяя водорослям улавливать максимум света для компенсации стресса.
Подобно «солнечной батарее», таллом в сухом состоянии защищён оранжевым «чехлом», а во влажном — снимает его, чтобы работать на полную мощность. Этот процесс не только оптическая иллюзия, но и индикатор жизнеспособности: в загрязнённых районах позеленение замедляется до 30–40 минут.
Симбиоз с бактериями
Современные исследования (2020-е гг.) раскрыли удивительный факт: помимо классического дуэта гриба и водоросли Trebouxia, таллом ксантории содержит сложное сообщество бактерий-симбионтов, выполняющих критические функции для выживания вида. Эти микроорганизмы образуют специализированные зоны: в растущих участках доминируют азотфиксирующие альфа-протеобактерии (Alphaproteobacteria) (до 45% биомассы), особенно представители рода Rhizobium, преобразующие атмосферный азот (N₂) и нитраты (NOₓ) в аммоний (NH₄⁺); в стареющих зонах — Acidobacteria, разлагающие полисахариды сердцевины и нейтрализующие тяжёлые металлы (Pb, Cd), связывая их в нерастворимые комплексы; а на поверхности Gammaproteobacteria формируют защитные биоплёнки против патогенов.
Механизм сотрудничества:
Гриб поставляет бактериям сахара (рибитол), а те взамен:
1) Детоксицируют загрязнения в городах,
2) Синтезируют антибиотики против патогенов,
3) Регулируют pH на экстремальных субстратах.
Бактериальное сообщество превращает ксанторию в «биореактор», способный колонизировать невозможные для других лишайников среды:
На токсичных серпентинитах (богатых Ni, Cr) бактерии Acidobacteria выделяют хелаторы (например, катехолы), связывающие тяжелые металлы в нетоксичные комплексы;
На кислых субстратах (pH<4.5) микобионт (грибной партнер) синтезирует атранорин – мощный природный буфер, а симбиотические бактерии (включая Actinobacteria) усиливают устойчивость, нейтрализуя локальную кислотность и подавляя конкурентов; При загрязнении реактивными формами азота (NOₓ, NO₃⁻, NO₂⁻) бактерии Rhizobium (и другие азотфиксаторы) преобразуют их в аммоний (NH₄⁺), доступный для питания фотобионта (водоросли).
Экология
Ксантория настенная это классический пример эврисубстратного лишайника, демонстрирующего феноменальную способность колонизировать чрезвычайно широкий спектр поверхностей как в природных, так и в антропогенных ландшафтах. Универсальность – ключевая черта её экологической стратегии. В природных условиях Ксантория предпочитает хорошо освещённые (гелиофильные) местообитания.
На коре деревьев она наиболее обильна на видах с нейтральной или слабощелочной корой (pH ~6.5-8.0), таких как ясень, клён, вяз и грецкий орех. На берёзах, чья кора часто имеет более кислую реакцию, она встречается реже и обычно в специфических микроместообитаниях – у основания стволов, на участках с повышенным стоком воды, несущей питательные вещества, или в условиях умеренного азотного загрязнения. На коре хвойных деревьев с сильнокислой корой встречается значительно реже, но не исключена.

На естественных скальных породах ксантория проявляет выраженное предпочтение к щелочным, богатым кальцием субстратам: известняку, доломиту, мрамору, мергелю и, в меньшей степени, базальту. Она активно заселяет трещины, карнизы и вертикальные поверхности этих пород, где меньше конкуренции со мхами и накапливается мелкозем. Кислые породы (граниты, кварциты, песчаники) в естественных условиях избегаются, если только они не выветрены или не покрыты кальцийсодержащими отложениями.
Однако истинный масштаб эврисубстратности ксантории раскрывается в антропогенной среде. Этот вид стал пионером и часто доминантом на самых разнообразных искусственных материалах:
Бетон и цемент: Один из наиболее частых и характерных колонистов, процветающий благодаря изначально высокому (щелочному) pH, который со временем снижается до оптимального диапазона. Шифер (асбоцементный): Широко распространена на старых крышах из-за щелочности и пористой поверхности материала.
Пластик: Способна медленно заселять старые, выветренные пластиковые поверхности (ПВХ, полиэтилен высокой плотности), где образуются микроповреждения и накапливается пыль, служащая якорной точкой.
Металл: Может расти на корродированных металлических поверхностях (железо, оцинкованная сталь), покрытых ржавчиной или минеральными отложениями.

Черепица: Часто встречается как на керамической, так и на бетонной черепице.
Стекло: Редко, но описаны случаи колонизации старого, матового или сильно загрязненного стекла.
Фундаментом этой уникальной эврисубстратности служит комплекс физиологических и симбиотических адаптаций. Ксантория обладает высокой толерантностью к широкому диапазону pH (от умеренно кислых ~4.5 до сильно щелочных >9.0 субстратов), ярко выраженной ксерофитностью (устойчивостью к высыханию) и гелиофильностью (потребностью в хорошем освещении). Критическую роль играет ее политолерантность – устойчивость к загрязнению воздуха, особенно к оксидам серы (SO₂) и оксидам азота (NOₓ), а также нитрофильность – способность использовать повышенное содержание азота в среде как преимущество. Эффективный симбиоз с водорослью Trebouxia и комплексом специализированных бактерий обеспечивает автотрофное питание, фиксацию атмосферного азота, детоксикацию тяжелых металлов и защиту от патогенов, что позволяет виду выживать и процветать на токсичных или физически сложных субстратах, недоступных для большинства других лишайников. Способность закрепляться в минимальных неровностях поверхности завершает картину ее вселенца.
Таким образом, сочетание широкой экологической амплитуды по ключевым факторам среды и уникальных симбиотических механизмов превращает Xanthoria parietina в космополитный вид, успешно захватывающий самые разнообразные ниши по всему миру, особенно в изменённых человеком ландшафтах, где она часто выступает индикатором определённых условий и фоновым видом.
Химическая идентификация и адаптации таллома
Химический состав Xanthoria parietina служит не только ключом к его безошибочной идентификации, но и раскрывает комплекс тонких адаптаций, обеспечивающих выживание в разнообразных условиях. Основу этой химической «сигнатуры» составляют специфические лишайниковые вещества (вторичные метаболиты), синтезируемые грибным партнером (микобионтом) и локализующиеся в определённых слоях таллома. Наиболее диагностически значимыми являются антрахинон парьетин (парьетиновая кислота, он же физцион) и депсидон атранорин.
Результаты стандартных химических проб с 20% растворами NaOH (гидроксид натрия) и KOH (гидроксид калия), предоставляют наглядное подтверждение химического профиля вида. При нанесении любого из этих реактивов на ярко-оранжевые апотеции (плодовые тела) и интенсивно окрашенные участки верхней поверхности таллома наблюдается мгновенная (в течение 1-3 секунд) и интенсивная вишнево-красная или пурпурно-красная реакция.

Эта яркая реакция (обозначаемая K+ красный, Na+ красный) однозначно указывает на присутствие парьетина и является главным диагностическим признаком Ксантории настенной, надежно отличающим её от сходных оранжевых лишайников (например, многих Caloplaca, дающих фиолетовую реакцию). Параллельно, обработка серовато-беловатого корового слоя теми же реактивами вызывает развитие слабого, но отчётливого лимонно-жёлтого или бледно-жёлтого окрашивания (K+ желтый, Na+ желтый). Эта реакция обусловлена наличием атранорина. Области сердцевины (медуллы) и неокрашенной нижней поверхности, как правило, значимых изменений не проявляют. Комбинация K+/Na+ красный на апотециях и K+/Na+ желтый на коре образует уникальный химический «отпечаток пальца» вида.
Экологическая роль лишайниковых веществ
Эти вторичные метаболиты – не просто маркеры, а активные участники экологической стратегии ксантории. Парьетин, концентрирующийся в верхних слоях и апотециях, выполняет критическую роль мощного фото- и УФ-протектора. Он эффективно поглощает губительное ультрафиолетовое излучение (особенно UV-B), защищая хлорофилл фотобионта и ДНК обоих симбионтов от повреждения, что жизненно необходимо для существования на открытых, солнечных субстратах. Его яркая оранжевая окраска также может служить предупреждающим сигналом для потенциальных травоядных. Атранорин, в свою очередь, является многофункциональным соединением. Он действует как эффективный антимикробный агент (антибактериальный и антифунгальный), подавляя рост патогенов и конкурентов на поверхности таллома и субстрата. Его буферные свойства помогают нейтрализовывать экстремальные значения pH (как в кислых, так и в щелочных условиях) в непосредственной близости от лишайника. Кроме того, атранорин обладает гидрофобным действием, способствуя отталкиванию излишков влаги и быстрому просыханию таллома после дождя или росы, что дополняет ксерофитные адаптации вида. Оба вещества, наряду с деятельностью бактерий-симбионтов, могут участвовать в связывании (хелатировании) токсичных тяжелых металлов, снижая их вредное воздействие.
Ареал распространения
Ксантория настенная (Xanthoria parietina) демонстрирует впечатляющую по масштабам географическую пластичность. Первичный (естественный) ареал вида охватывает всю Голарктическую область – обширный биогеографический регион, включающий Европу (от Скандинавии до Средиземноморья), значительную часть Азии (от Ближнего Востока и Сибири до Дальнего Востока России, Японии и Корейского полуострова) и Северную Америку (от Аляски и Канады через США до горных районов Мексики). В пределах этого гигантского пространства вид встречается преимущественно в умеренном климатическом поясе, но также проникает в субарктические (южные тундры) и субтропические зоны (например, в горных системах Средиземноморья или Кавказа).
Уникальность ксантории заключается в её стремительной экспансии в Южное полушарие, превратившей вид в подлинного космополита антропогенных ландшафтов. Эта экспансия – прямое следствие человеческой деятельности, начавшейся с эпохи Великих географических открытий и усилившейся в XX-XXI веках. Сегодня вид надёжно натурализован и широко распространён:
В Австралии и Новой Зеландии, где он обильно покрывает деревья в сельскохозяйственных районах, бетонные сооружения в городах и естественные скалы в прибрежных зонах.
В Южной Африке, особенно в регионах с зимними осадками (например, Западно-Капская провинция), на фруктовых фермах и в урбанизированных зонах.
В Южной Америке – в умеренных районах Чили и Аргентины (Патагония, Анды), а также в антропогенных местообитаниях Бразилии и Уругвая.
На многочисленных океанических островах, включая Гавайи, Фолклендские острова, остров Святой Елены и даже некоторые субантарктические острова (например, Кергелен), куда он был занесён человеком.
Съедобность
Несмотря на яркую, почти «аппетитную» внешность, Ксантория настенная не является съедобной для человека и не относится к лекарственным растениям в официальной медицине. Причин для этого несколько:
1. Токсичность вторичных метаболитов: Ключевые лишайниковые вещества ксантории – парьетин (физцион) и атранорин – обладают выраженным раздражающим действием на желудочно-кишечный тракт человека. Атранорин, придающий таллому характерную горечь, в высоких дозах может вызывать тошноту, рвоту и диарею. Хотя системные исследования токсичности именно парьетина для млекопитающих ограничены, его принадлежность к классу антрахинонов (многие из которых известны слабительным или раздражающим эффектом) делает употребление лишайника в пищу потенциально опасным.
2. Накопление загрязнителей: Как активный биоаккумулятор, Ксантория эффективно поглощает из воздуха и субстрата тяжелые металлы (кадмий, свинец, цинк, медь) и другие токсичные соединения (включая радионуклиды). Особенно высоки концентрации загрязнителей в талломах, растущих в городах, вдоль дорог или в промышленных зонах. Употребление такого «сырья» чревато серьезным отравлением.
3. Низкая питательная ценность и плохая усвояемость: Таллом лишайника состоит в основном из трудноусвояемых грибных полисахаридов (лихенина). Человеческий организм не обладает ферментами, необходимыми для эффективного расщепления этих специфических углеводов и хитиноподобных компонентов клеточных стенок гриба. Это делает лишайник малопитательным и трудноперевариваемым продуктом.
Использование в народной медицине
Официальная (доказательная) медицина не использует Xanthoria parietina в качестве лекарственного сырья. Нет утверждённых препаратов на её основе, и клинические исследования, подтверждающие безопасность и эффективность для человека, отсутствуют.
В некоторых регионах (например, отдельные традиции в Европе и Азии) Ксантория настенная исторически могла упоминаться в народной медицине, обычно в контексте:
«Очищения» печени и желчегонного действия (ирония, учитывая потенциальную гепатотоксичность атранорина!).
Наружного применения в качестве антисептика (благодаря доказанным in vitro антимикробным свойствам атранорина и парьетина) для обработки ран или кожных заболеваний.
Лечения респираторных заболеваний.
Однако использовать эти рецепты категорически не рекомендуется! Концентрация активных (и токсичных) веществ в талломе непредсказуема, зависит от места сбора и экологических условий. Риск отравления тяжёлыми металлами или собственными метаболитами лишайника, а также отсутствие стандартизации и доказанной пользы многократно перевешивают любые гипотетические преимущества. Самолечение лишайниками опасно для здоровья.
Сходные виды
Прежде всего обратим внимание на род Xanthoria (Ксантория), объединяющий около 19 видов накипных и листоватых лишайников, характерных яркой жёлто-оранжевой окраской таллома, обусловленной пигментом париетином. Многие виды визуально похожи, особенно в полевых условиях, что требует внимания к деталям субстрата, морфологии и распространения.
Ксантория известняковая (Xanthoria calcicola)
Оба вида образуют крупные (3–10 см) оранжево-жёлтые розетки с широкими лопастями (до 5 мм), часто с апотециями по центру.
Ключевые отличия:
Лопасти более жёсткие и матовые, с приподнятыми краями. Нижняя поверхность светло-серая. При нанесении KOH таллом краснеет интенсивнее, чем у Ксантории настенной из-за высокой концентрации париетина.
Ксантория известняковая строгий кальцефил — встречается исключительно на карбонатных породах: известняк, доломит, мрамор, старый бетон или строительный раствор. На коре деревьев не развивается.
Редкий вид с дизъюнктивным ареалом (Кавказ, Южный Урал, Алтай). Занесён в Красные книги Липецкой и Свердловской областей. Приурочен к старым известняковым карьерам или скальным выходам в степной зоне.
Калоплака наскальная (Caloplaca saxicola)
Хотя оба вида – жёлто-оранжевого цвета, часто растущие бок о бок на известняковых скалах, старых стенах или бетонных сооружениях, их легко отличить по ряду ключевых признаков, касающихся формы таллома, окраски, структуры плодовых тел и химических свойств.
У Ксантории настенной таллом листоватый. Состоит из отчетливо выраженных, широких, плоских, округлых лопастей.
У Калоплаки наскальной накипной (корковидный) таллом. Ее слоевище плотно прирастает к субстрату всей своей нижней поверхностью и не образует приподнятых лопастей. Лишь край слоевища образует ареол из радиально направленных тесно скученных, выпуклых или угловатых чешуек.
Апотеции у Калопаки крайне многочисленны, мелкие и часто настолько плотно покрывают поверхность таллома, что почти скрывают его. Диск апотеция почти всегда более яркий и насыщенный, чем таллом. Окрашен в оранжево-красные или густо-оранжевые цвета. Эта «пёстрая» картина из множества красных точек на желтом фоне очень характерна.
Проба с KOH на апотециях или талломе Калоплаки наскальной даёт совершенно иную реакцию чем у Ксантории – яркое фиолетовое или пурпурное окрашивание (K+ фиолетовый). Иногда оттенок может быть красно-фиолетовым. Эта реакция обусловлена другими антрахинонами (вероятно, эмодином или родственными соединениями). Отсутствие интенсивной чисто красной реакции KOH надёжно исключает Ксанторию.
Калопака оранжевая (Caloplaca aurantia)
Встречается повсеместно в средней полосе Восточной Европы и России. Визуальным сходством с Ксанторией настенной можно назвать ярко оранжевый таллом. Однако эти виды легко различимы по типу таллома. Если у ксантории он листоватый с приподнимающимися лопастями, то у Калопаки плотно приросший корковый таллом, не формирующий лопастей. Лишь по краю виден ареол из вздутых радиально направленных чешуек. Кроме того апотеции у Xanthoria parietina крупнее, часто с сохраняющимся слоевищным краем, а у Caloplaca aurantia – мельче, обычно выпуклые и без края. Наиболее надежным экологическим различием является предпочтение субстрата: Калопака оранжевая – облигатный кальцефил, строго приуроченный к известнякам, бетону и строительному раствору, в то время как Xanthoria parietina растёт на самых разнообразных субстратах, включая кору деревьев и силикатные породы, не требуя наличия кальция. Микроскопически виды различаются строением спор (одноклеточные у Ксантории, двуклеточные у Калопаки).
В 2007 году Европейское космическое агентство (ESA) провело дерзкий эксперимент: образцы Xanthoria parietina отправили в имитатор марсианской среды с убийственными параметрами:
— Вакуум глубже космического,
— Температура -196°C (жидкий азот!),
— Жёсткое УФ-излучение, в десятки раз сильнее земного.
Результат ошеломил учёных: после 14 дней пытки лишайник не просто выжил – он продолжал фотосинтезировать после возвращения в земные условия! Секрет кроется в париетине: пигмент образовал щит, поглотивший 99% смертоносного излучения и стабилизировавший клеточные структуры.
Этот опыт доказал: даже сложные организмы способны пережить межпланетный перелёт. Сегодня Ксантория – ключевой модельный объект в разработке биозащиты для колонистов Марса.
Народные названия
- Золотянка
- Стенная золотнянка
- Дворовый лишайник
- Солнечник
- Кирпичный мох
- Лишайник-апельсин
- Морское солнышко
- Асфальтовый цветок
- Птичья карамель




Фото гриба Ксантория настенная из вопросов в распознавании:






















Ксантория настенная упоминается в отчетах:
- Ксантория настенная на разных поверхностях 22 апреля 5 6












Ого! Супер! В это надо вникать без спешки. Фото роскошные! Особенно понравилось на железной сетке. Вот бы лобавить на бетоне и на стекле!
Спасибо за добрые слова
Вы правы, что на бетоне можно добавить. Надо будет со временем «хваст» сделать. Так хоть опосредованно добавится.
А на стекле я лично ещё не видел. 🤷
А в той пачке, которую Вы присылали, есть фото на бетоне? Если есть, могу добавить в статью.
Хваст — отличная идея!
Вроде бы не было. Как то об этом был не подумал.
Впрочем это не проблема могу сфотографировать и позже прислать.
Сфотографируйте и сделайте Хваст. Будет замечательно.
Хорошо 👍, так и сделаю.